
Texte et illustrations : Joseph HEMMERLÉ
Avec l’aimable autorisation de Joseph HEMMERLÉ et de son éditeur – Article paru initialement dans la revue Fruits et Abeilles (N° d’avril 2021, pages 135 à 137 et mai 2021, pages 155 à 157)
Joseph HEMMERLÉ était présent lors de notre journée technique du 11 Mars 2023 pour animer une conférence passionnante sur les mécanismes de l’essaimage.
Ses différents ouvrages sur le Pollen, le Miel et la Santé des abeilles sont disponibles sur le site de son éditeur Les Editions TERRAN
Si un apiculteur a des cheveux gris, il est probable que l’essaimage de ses protégées y soit pour quelque chose… Trêve de plaisanterie, il faut reconnaitre que malgré les efforts de l’Homme pour domestiquer l’abeille mellifère, le statut de l’insecte demeure ambivalent. Une colonie d’abeilles mellifères peut être facilement récupérée et exploitée par un apiculteur ; mais une colonie d’abeilles « domestiques » peut tout aussi facilement reprendre sa liberté et survivre en totale autosuffisance. Et, tout compte fait, malgré leurs finalités apparemment paradoxales, essaimage et « domestication » de l’abeille sont intimement liés. En quelque sorte, l’essaimage et la récupération d’essaim s’apparentent au jeu du chat et de la souris. Quel apiculteur ne se souvient pas avec fierté, et parfois émotion, de sa première cueillette d’un essaim ? Combien d’aventures, parfois rocambolesques, se racontent sur le sujet ?
Le terme « essaimage » est beaucoup trop réducteur pour couvrir l’ensemble des mécanismes qui le composent: l’essaimage naturel n’est pas un événement, c’est un processus Ce mot englobe une cascade d’événements complexes savamment intriqués. Parmi eux, quatre phénomènes, très étudiés mais gardant toujours des parts de mystères, intriguent la communauté scientifique et bien entendu les apiculteurs :
- Les signes annonciateurs d’un essaimage ;
- Les mécanismes de prises de décision d’un essaim ;
- Les « chants » des reines ;
- Les étapes menant à la restauration de la monogynie après l’essaimage.
Ce qui peut déclencher un essaimage
La mise en route d’un processus d’essaimage procède d’une combinaison de facteurs déclenchants, dont :
- Une intense activité d’élevage ;
- Un rapide développement de la colonie ;
- Une congestion du nid à couvain ;
- Une forte augmentation du nombre de jeunes abeilles ;
- Une diffusion insuffisante ou inhomogène de phéromone royale.
En ce qui concerne ce dernier point, rappelons que les phéromones sont des molécules volatiles qui agissent dans un rayon intra-spécifique. Ces médiateurs chimiques engendrent des réponses physiologiques ou comportementales chez les congénères. Ainsi, la phéromone mandibulaire de la reine, composée de plusieurs substances, joue deux rôles majeurs dans le contexte de l’essaimage :
- Elle inhibe l’instinct d’étirement de cellules royales chez les ouvrières ;
- Elle assure la cohésion de groupe de l’essaim en vol d’abord, puis dans la grappe temporaire ensuite.
Les causes d’une insuffisance en phéromone royale dans la ruche peuvent être de deux ordres : soit la reine âgée ne produit plus assez de ces messages chimiques, soit la diffusion de ces molécules volatiles n’est plus assurée dans tout le volume de l’abri.
Par ailleurs, il faut savoir que la propension à l’essaimage varie aussi selon les races d’abeilles.
Chronologie des préparatifs d’essaimage
Pour commencer la colonie d’abeilles donne naissance à deux clans : ceux qui vont partir et ceux qui vont rester. Une fois que la fièvre d’essaimage est enclenchée, c’est « battez tambour et sonnez trompettes ! ». Il est alors très difficile pour l’apiculteur d’arrêter le processus. La chronologie des principaux événements qui jalonnent la préparation d’un essaimage peuvent être résumés comme suit :
1) Au moins quinze jours avant le départ de l’essaim, la reine est mise à la diète et les ouvrières édifient une douzaine de cellules royales environ dans des zones périphériques des rayons.
2) Environ dix jours avant l’essaimage, les ouvrières concernées commencent à se gorger de miel (jusqu’à 35 mg par ouvrière). De plus, des sons se font entendre. La fréquence acoustique initiale de 100 à 200 Hz atteint finalement 200 à 250 Hz. Le nombre de séquences sonores, d’une durée moyenne d’une seconde, émises par les ouvrières augmente de jour en jour. Il s’agit là d’une stratégie de mobilisation des acteurs du processus d’essaimage.
3) Environ huit jours avant le départ, on note une diminution de la ponte de la reine.
4) Finalement, dans l’heure qui précède le départ de l’essaim, des ouvrières prêtes à s’en aller pratiquent une « marche bourdonnante » qui s’intensifie jusqu’à l’envol d’un ensemble impressionnant d’abeilles (Photo illustrant l’article – Crédit : Joseph HEMMERLÉ).
L’envol de l’essaim
Le départ d’un essaim primaire se produit après l’operculation d’au moins une première cellule royale. Les essaimages sont fréquents aux alentours du mois de mai et se produisent généralement entre 10 h et 16 h, par temps ensoleillé. Toutefois, le départ du groupe d’abeilles est conditionné par la météo.

Une fois que toutes les conditions nécessaires sont réunies, la reine, entourée d’un grand nombre d’ouvrières, quitte la ruche dans un bruissement enflammé caractéristique. La nuée d’insectes s’agrège en une grappe accrochée à un support situé à un jet de pierre de l’ancienne demeure (figure 2). Deux raisons expliquent la courte distance parcourue par l’essaim : premièrement la reine manque d’entrainement au vol et, deuxièmement, le groupe d’abeilles qui a pris le large ne sait pas où aller. La configuration de l’essaim, suspendu transitoirement à un support de fortune, se structure différemment selon les conditions météorologiques :
- Par temps froid et/ou pluvieux, la grappe d’abeilles se contracte et s’entoure d’un manteau multicouche d’insectes serrés les uns contre les autres. Cette conformation conduit à un gradient de température, décroissant du centre vers l’extérieur, dans l’amas temporaire d’abeilles.
- Par temps chaud, la configuration de l’amas d’abeille cherche à éviter les situations d’hyperthermie. C’est pourquoi la grappe est plus dilatée et dépourvue d’un manteau compact d’insectes. De plus, l’interconnexion d’espaces laissés vides forme dans la grappe des conduits de ventilation qui facilitent la régulation thermique.
La composition d’un essaim
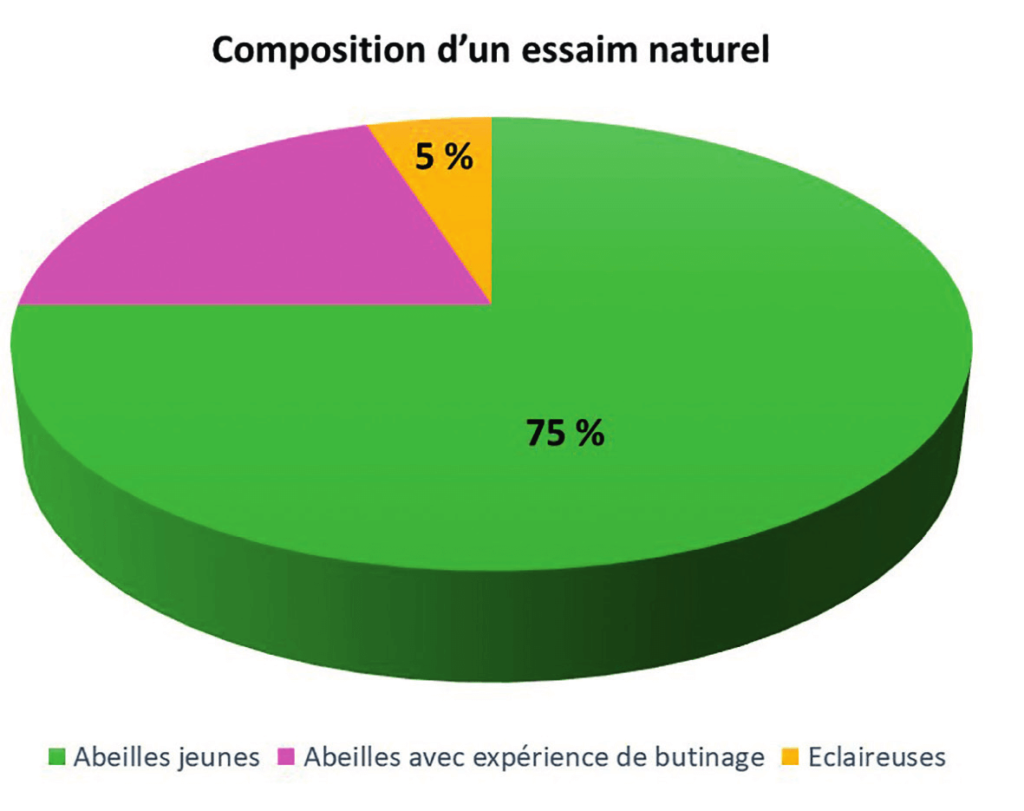
Un essaim primaire compte communément entre 10 000 et 20 000 ouvrières et le poids de sa reine avoisine les 200 mg. La phéromone royale assure l’agrégation de la grappe d’abeilles. La majorité (environ trois quarts) des abeilles qui constituent l’essaim sont de jeunes abeilles ayant moins de dix jours. Et pour cause : elles doivent être capable de construire le nouveau nid, soit près d’un mètre carré de rayons de cire, et pouvoir conduire ex nihilo un élevage complet de couvain. Le quart restant d’abeilles, principalement localisées à la périphérie de la grappe provisoirement suspendue, sont des insectes d’âge moyen, qui ont déjà une certaine expérience de butinage. Et c’est de ce dernier groupe d’ouvrières que sont issues les « éclaireuses » ; ces abeilles qui vont prospecter pour un nouveau site de nidification. Près de 5 % de la population de l’essaim font office d’éclaireuses (figure 3).
En route vers le nouveau nid…
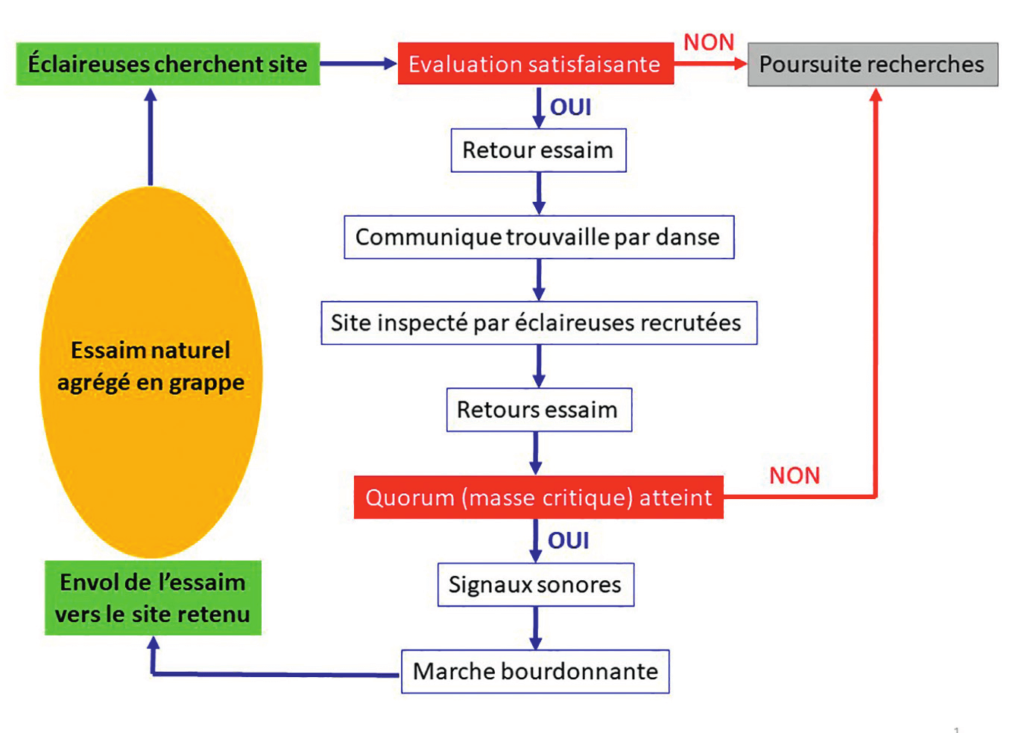
Le logigramme de la figure 4 décline les différentes étapes du processus de sédentarisation d’un essaim. Dans ce schéma d’exécution, trois séquences sont cruciales :
- 1) Trouver un site de nidification ;
- 2) Convaincre le plus grand nombre d’éclaireuses ;
- 3) Emmener l’essaim jusqu’au site retenu.
Pour y voir plus clair, décortiquons chacune de ces étapes
1) Trouver un site de nidification
Pour commencer, des éclaireuses partent en reconnaissance, dans toutes les directions, à la recherche d’abris potentiels de nidification. Lorsqu’elles repèrent un site en adéquation avec les besoins de la future colonie, ils reviennent vers l’essaim et communiquent leur trouvaille aux abeilles en attente via une danse pratiquée à la surface de la grappe temporaire. Les danses sont un mode de communication commun pour les abeilles. C’est un langage simple, mais adapté, suffisant et efficace. En réponse à cette danse de renseignement, d’autres éclaireuses iront vérifier la conformité du site proposé ; et si elles aussi adhèrent au choix de site, elles reviennent sur l’essaim pour constituer un ensemble grandissant d’éclaireuses qui pratiquent la même danse informative.
2) Convaincre le plus grand nombre d’éclaireuses
Au fur et à mesure des prospections, il se forme plusieurs groupes d’éclaireuses qui tous essayent de dicter leur avis quant à un choix de site. Dans cet apparent chaos, le verdict final appartiendra aux plus convaincants !
Les interactions se déroulent en deux temps et à deux niveaux : d’abord exclusivement entre les éclaireuses et ensuite entre un groupe d’éclaireuses et les abeilles qui forment la masse de la grappe. En effet, les ouvrières ne délibèrent pas pour choisir la meilleure des options de nouvel abri défendues par les différents groupes d’éclaireuses. Comment le pourraient elles, puisqu’il s’agit d’individus jeunes et inexpérimentés qui, de surcroit, n’ont pas visité les différentes solutions de relogement ? La première phase spatio-temporelle du processus décisionnel se déroule au niveau des éclaireuses et à la surface de la grappe. En l’état actuel des connaissances, il apparait que c’est un effet de seuil, plutôt qu’un effet de consensus, qui déclenche les préparatifs d’envol de l’essaim. Pour convaincre, la stimulation induite par un groupe d’éclaireuses militant pour un même endroit doit franchir un seuil de participants. En d’autres termes, les abeilles se mobilisent pour l’envol dès qu’un nombre déterminant (quorum) d’éclaireuses, mais pas toutes, se sont accordées sur un choix de site de nidification. Il semblerait que ce mode opératoire soit le plus efficace pour permettre à un aussi grand nombre d’individus de s’entendre sur une destination dans un délai compatible avec l’autonomie énergétique de l’essaim.
Lorsqu’un quorum est atteint, les éclaireuses du groupe s’étant imposé émettent des signaux sonores (chants) de fréquences croissantes, qui engendrent une réponse enfiévrée de la grappe d’abeilles. On assiste à une acceptation d’adhésion de l’essaim, alors même que la majorité des insectes n’ait pas participé à la délibération. La mise en route du processus de stimulation démarre entre une et deux heures avant l’envol de l’essaim. En réponse au signal vibro-acoustique, les jeunes ouvrières de l’essaim, qui s’économisaient jusque-là, échauffent leurs muscles de vol. Ensuite, dans l’heure qui précèdent la dissociation de la grappe, les abeilles excitatrices remplacent progressivement les chants par des « marches bourdonnantes ». Finalement, l’excitation collective atteint son apogée et l’agrégat d’abeilles se défait. Les abeilles débordent d’enthousiasme ! Elles ne savent pas où aller ? Qu’à cela ne tienne : les éclaireuses aguerries les guideront !
3) Emmener l’essaim jusqu’au site retenu
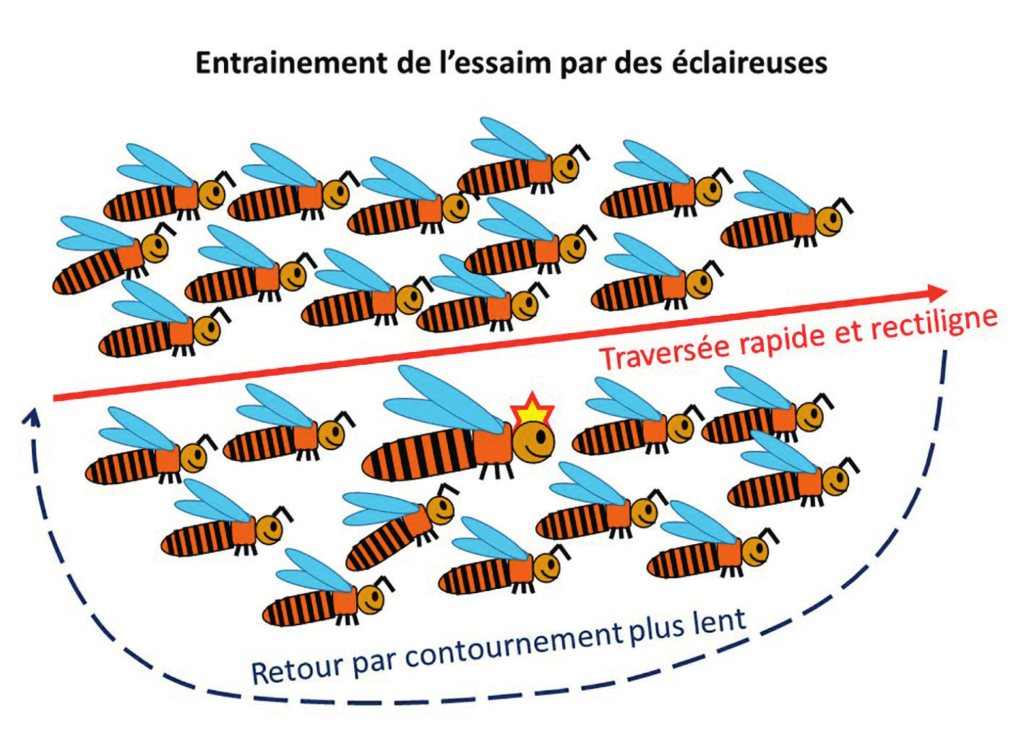
Comment les éclaireuses, qui représentent moins de 5 % de la population de l’essaim et qui ont connaissance du nouveau site de nidification, procèdent-elles pour guider la nuée d’abeilles, volant à quelques mètres au-dessus du sol, jusqu’à la destination providentielle ? La migration de ce nuage d’abeilles uni par l’action de la phéromone royale a tout d’un « entrainement directif ».
Des enregistrements réalisés avec des caméras de haute définition ont révélé les secrets de cet entrainement directif (figure 5). Les éclaireuses traversent avec un vol rapide et rectiligne l’essaim en mouvement. Arrivées au-devant de la nuée d’abeilles frétillantes, les éclaireuses contournent l’essaim, avec un vol plus lent, jusqu’à la « queue » du nuage d’insectes, puis réitèrent une traversée à grande vitesse. Ces séquences de vol rapide et rectiligne à travers l’essaim suivies d’un contournement plus lent de la masse d’insectes en mouvement se répètent jusqu’à ce que les abeilles atteignent le nouveau refuge. Et ça marche !
Les mécanismes de l’essaimage naturel – Partie 2
Après nous être intéressés aux abeilles qui sont parties, celles de l’essaim, le moment est venu de nous pencher sur le devenir des ouvrières et reines qui sont restées dans la ruche source.
La colonie d’origine, celle d’où est parti l’essaim, se retrouve avec moins d’ouvrières, mais aussi… avec un surplus de reines en gestation. La situation est similaire à celle d’une campagne présidentielle : plusieurs candidates brigueront l’unique position royale. Sauf que, chez les abeilles, la course au poste suprême est un peu plus « virile » que dans une démocratie humaine.
Restaurer la monogynie
Le défi de la colonie d’origine est de retrouver le monde d’avant : une seule reine (monogynie) à la tête d’un peuple uni. Ça sonne bien, mais la méthode pour y parvenir fait froid dans le dos ! En effet, comme les abeilles avaient élevé plusieurs larves royales, l’élimination des reines surnuméraires est donc consubstantielle à la restauration de la monogynie. L’héritage du nid maternel devra donc se conquérir.
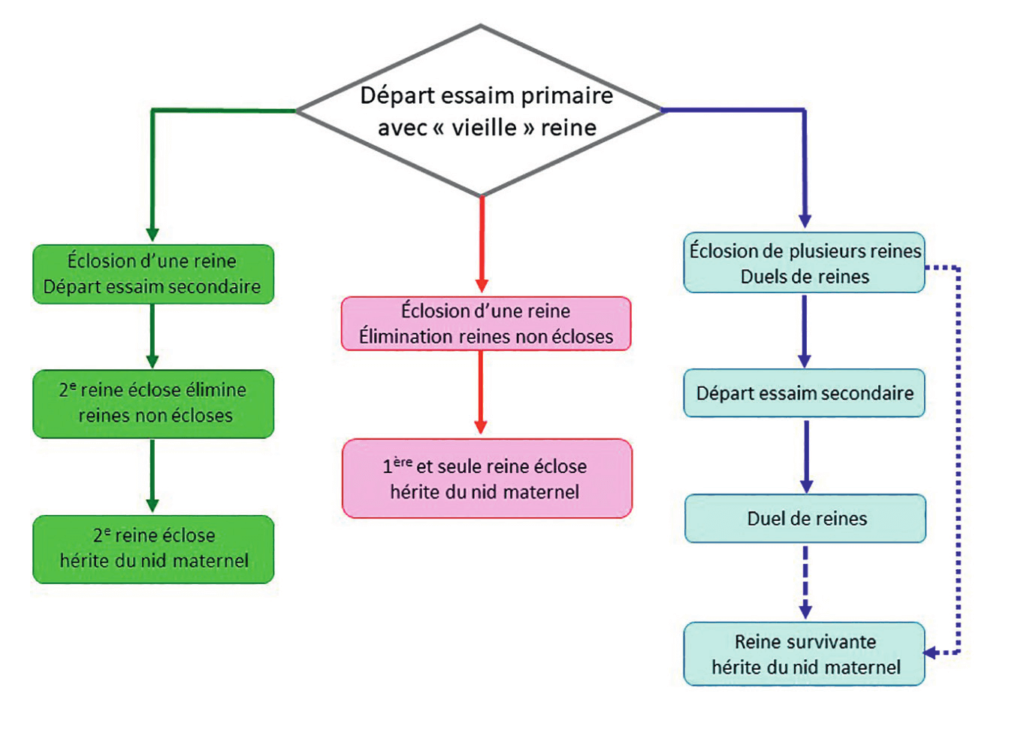
La période d’« épuration royale » débute peu après l’émergence de la première reine. Trois scénarios d’élimination de reines sont envisageables :
- La mort d’une reine suite à un duel avec une autre reine soeur ;
- L’assassinat de reines avant leur émergence par une reine soeur déjà éclose ;
- La fuite d’une reine avec un essaim.
Les duels de reines sont ponctués par des étapes de fuites et de combats. Dans les faits, les reines passent le plus clair de leur temps à patrouiller sur les rayons de la ruche. Des observations révèlent que les reines écloses se croisent toutes les 45 minutes environ. Lorsqu’elles passent à quelques centimètres l’une de l’autre, il ne se passe pas grand-chose : elles se contentent de se toiser et de se jauger. Il arrive quand même qu’une prétendante un peu plus hardie asperge sa rivale de matières fécales ! En revanche, lorsque deux reines tombent nez à nez, elles s’agrippent avec les pattes et les mandibules, se battent et tentent de se piquer mutuellement. Ces bagarres durent entre quelques secondes et quelques minutes. Les pugilats se concluent généralement par la blessure ou la mort de l’une des deux combattantes.
Le synoptique de la figure 6 schématise, dans les grandes lignes, les principales voies menant à la restauration de la monogynie dans la ruche source. Ce tableau souligne la remarquable plasticité d’une société d’abeilles pour s’adapter aux situations les plus diverses d’un remérage naturel. Le plus surprenant dans cette affaire, c’est que ces « éliminations » de reines sont orchestrées par des « chants » à deux voix : des « tut » et des « couac »…
Des chants orchestrent le retour à la monogynie
Tout apiculteur ayant de la bouteille a déjà entendu les fameux « tut » des reines vierges candidates à la fonction maternelle. Mais il faut savoir qu’il ne s’agit là que d’une partie des dialogues qui parcourent la ruche. Explications :
1) Une première reine vierge éclos entre quelques heures et plusieurs jours après le départ de l’essaim (primaire). Elle émet des « tut » pour claironner son émergence. Ces signaux polysyllabiques, qui durent généralement moins de 10 secondes, ont une fréquence se situant entre 400 et 500 Hz. Pour pouvoir se faire une idée de la hauteur de ce son, notons que la fréquence fondamentale donnée par un diapason standardisé, la note La3 (3e octave), est de 440 Hz. Ces séquences de « tut » ont pour effet d’inhiber l’effort d’éclosion des reines soeurs encore enfermées dans leurs cellules. On peut observer qu’en réponse aux « tut », des ouvrières colmatent des cellules royales en voie d’ouverture (figure 7).
2) De leur côté, les reines matures toujours confinées dans leurs cellules répondent aux « tut » de la reine éclose avec des sons plus graves, des « couac » dont les fréquences oscillent entre 300 et 350 Hz. Ces réponses systématiques, après quelques secondes seulement, sont des répétitions d’une vingtaine de « couac » qui durent un tiers de seconde chacun. Les « couac » émis en écho aux « tut » signalent à la reine éclose, ainsi qu’aux ouvrières, la présence d’une ou plusieurs reines non encore écloses.

Bien que ce langage rudimentaire de « tut » et de « couac » nous paraisse abscons, il parle pourtant aux abeilles. Pour autant, l’apiculteur qui entend pour la première fois des « tut » royaux est rempli d’émotions et a l’impression d’intégrer quelque peu le monde des abeilles.
Des essaimages multiples
Il se peut qu’un essaim secondaire quitte la ruche source après la (ou les) première(s) émergence(s) de reine(s). Mais ces répliques d’essaimage comptent moins d’abeilles que l’essaim primaire. Dans les régions tempérées, la probabilité qu’un essaim secondaire survive à l’hiver suivant l’essaimage est faible. Il y a principalement deux raisons à cela :
- Tout d’abord, un essaim secondaire dispose de moins d’éclaireuses que l’essaim primaire : ce qui diminue la probabilité de trouver un abri adéquat dans un temps raisonnable.
- Ensuite, un essaim secondaire doit compter sur un nombre restreint d’ouvrières pour construire le nouveau nid, élever le couvain et faire des réserves pour l’hiver.
Néanmoins, vu depuis la ruche source, la fuite d’une reine vierge dans le sillage d’un essaim secondaire s’apparente à l’élimination d’une reine.
Plusieurs modes de communications
Une colonie d’abeilles en phase d’essaimage utilise des signaux chimiques et physiques pour coordonner les actions de plusieurs dizaines de milliers d’individus. La phéromone mandibulaire royale, composée de plusieurs molécules, intervient dans les mécanismes de régulation d’un essaimage. Les phéromones sont des molécules volatiles qui permettent une communication intraspécifique. Ces substances chimiques engendrent des réponses comportementales ou physiologiques chez des individus conspécifiques. Par ailleurs, des sons typiques, les « chants » des reines notamment, accompagnent certaines étapes d’un processus d’essaimage. À l’approche d’un essaimage, on note déjà que le niveau et les fréquences sonores augmentent dans la colonie. Des fréquences sonores plus élevées, pouvant atteindre 600 Hz, apparaissent parmi le bruit de fond ordinaire (100 – 300 Hz) de la ruche. Bien entendu, l’apiculteur ne peut pas détecter les phéromones émises par l’insecte, mais, par contre, il peut entendre les signaux sonores des abeilles, puisque l’humain entend des sons dans une gamme de fréquences située grosso modo entre 20 et 20 000 Hz.
Deux types de signaux vibratoires
Un insecte peut produire des signaux vibratoires dans un support en le frappant avec une partie de son corps. À cet égard, on connait chez l’abeille le signal de vibration abdominale dorso-ventrale qui stimule les diverses activités coopératives. Les différentes classes d’âges réagissent à ces vibrations selon leurs fonctions spécifiques : les nourrices s’activent dans leur mission d’élevage, et les butineuses se pressent vers les fleurs pour approvisionner la colonie. Ce signal permet aussi de synchroniser des processus d’essaimage. La vibration abdominale dorso-ventrale est perçue par les abeilles, mais demeure inaudible pour nous parce que sa fréquence, comprise entre 10 et 20 Hz, n’entre pas dans notre champ auditif. Le rayon de cire est le porteur de cette onde vibratoire, dont le rayonnement se diffuse surtout à l’échelle du cadre. L’abeille capte des signaux vibratoires véhiculés par le rayon de cire grâce à l’organe subgénual qui se trouve dans la partie proximale du tibia de ses six pattes (figure 8).

Mais en quoi consistent exactement les fameux « chants » des reines ? Les bruits que nous, humains, pouvons entendre dans une ruche ne sont pas nécessairement des signaux informatifs pour les abeilles. Pourtant, des sons plus précis, dont les fréquences se situent généralement entre 300 et 500 Hz, peuvent bel et bien être distingués dans les ruches.
Contrairement aux criquets et aux sauterelles, les abeilles ne sont pas équipées d’organes spécifiquement dévolus à la production de sons. Toutefois, l’abeille a la capacité de produire des sons grâce à la trémulation des muscles alaires, qui engendre des oscillations du compartiment thoracique. Des sons aéroportés peuvent être produits lorsque les ailes de l’insecte sont complètement repliées sur son dos ; ce qui est faisable puisque les muscles thoraciques ne sont pas directement attachés aux ailes. Les ailes transforment alors l’énergie mécanique de la trémulation en énergie vibratoire aérienne. Et si l’on parle du « chant » des reines, c’est simplement parce que la composante du signal que perçoit l’humain est d’ordre acoustique.
Qu’il s’agisse donc des « tut » ou des « couac », il s’agit avant tout d’une communication à base de signaux vibro-acoustiques. Les vibrations thoraciques des reines peuvent être transmises au rayon par contact direct du thorax sur la cire ou à travers les pattes (figures 9).
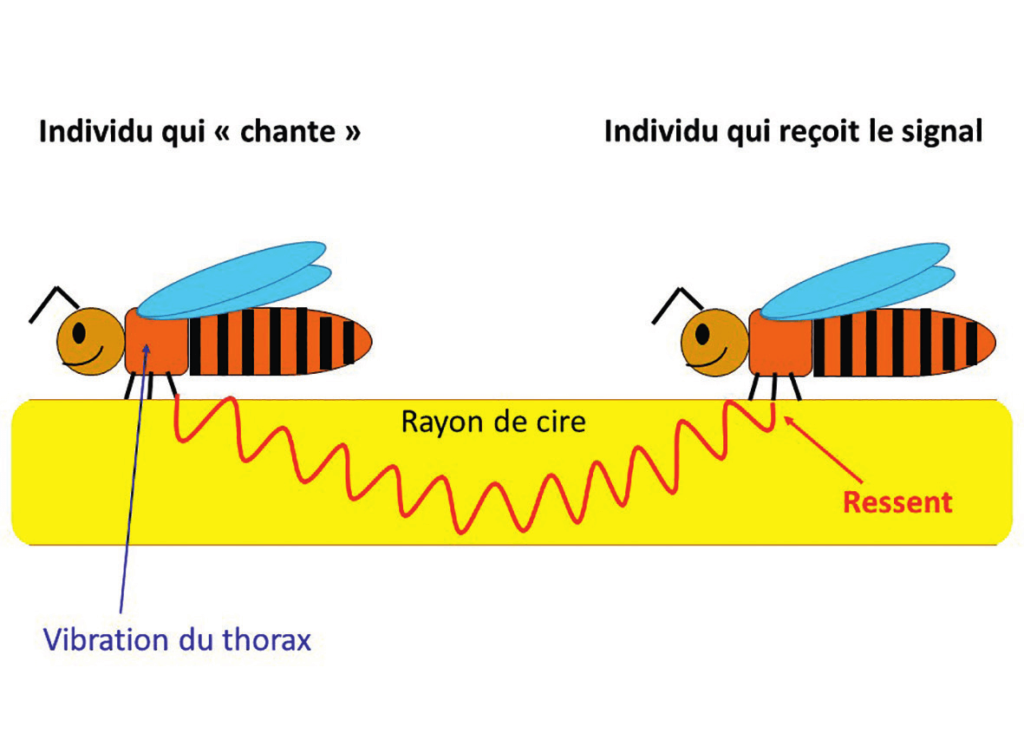
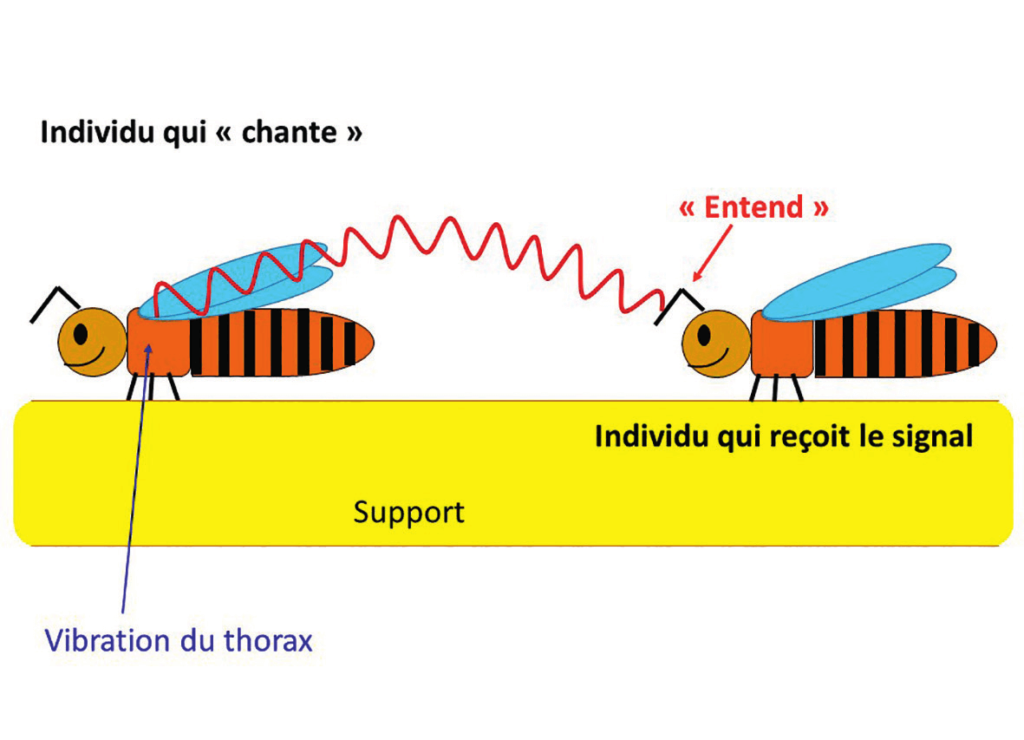
La composante mécanique du signal est interceptée par les pattes (organe subgénual) des congénères. En revanche, les sons issus de la trémulation des muscles alaires, qui sont véhiculés par l’air (figure 10), sont détectés par l’organe de Johnston situé dans le pédicelle des antennes (figure 11).
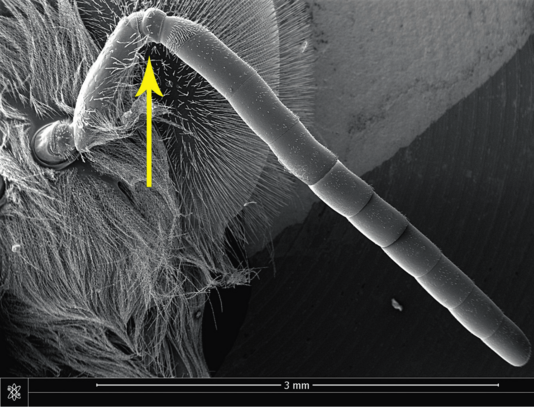
Ce capteur sensoriel peut détecter des déplacements de quelques dizaines de nanomètres seulement du bout du flagelle de l’antenne. La sensibilité du capteur est maximale pour des ondes acoustiques avoisinant les 350 Hz. Les « couac » émis par les reines encore prisonnières de leurs cellules correspondent à un son assourdi, pauvre en harmoniques. Nous l’aurons compris, comme nous ne percevons que les signaux royaux véhiculés par l’air, les « tut » et les « couac », il y a tout un pan de la communication vibratoire des abeilles qui nous échappe…
L’installation d’une unique nouvelle reine marquera la fin de l’épisode d’essaimage, et son retour du vol nuptial ouvrira la voie à la naissance d’une colonie renouvelée.
Les illustrations datant d’avant le Moyen Âge, qui décrivent l’apiculture, montrent que seules deux interventions rythmaient l’activité d’un éleveur d’abeilles : la récolte du miel et la capture d’essaims. C’est dire combien l’essaimage a de tout temps marqué les esprits, et pas seulement ceux des apiculteurs. Les quelques points que nous avons considérés ne sont bien sûr qu’un pâle reflet, forcément incomplet, d’une dynamique d’essaimage. Les différentes étapes de ce processus, comme la recherche d’un nouveau site de nidification, la transformation d’une « proposition » d’abri des éclaireuses en un « projet » de relogement accepté par l’ensemble des abeilles de l’essaim, la restauration de la monogynie et les chants singuliers des reines gardent toujours leurs parts de mystères, tellement ces prouesses biologiques et éthologiques sont étonnantes !
Références bibliographiques :
- Camazine S. et co-auteurs. House-hunting by honey bee swarms : collective decisions and individual behaviors. Insectes Sociaux 46 : 348-360, 1999.
- Fell R. et co-auteurs. The seasonal cycle of swarming in honeybees. Journal of apicultural Research 16(4): 170-173, 1977.
- Grozinger C. et co-auteurs. From molecules to societies: mechanisms regulating swarming behavior in honey bees (Apis spp.). Apidologie 45 :327-346, 2013.
- Heinrich B. The mechanisms and energetics of honeybee swarm temperature regulation. Journal of Experimental Biology 91 : 25-55, 1981.
- Schultz K. et co-auteurs. The mechanism of flight guidance in swarms : subtle guides or streaker bees ? The Journal of Experimental Biology 211 : 3287-3295, 2008.
- Seeley T. et Visscher P. Choosing a home : how the scouts in a honey bee swarm perceive the competition of their group decision making. Behavioral Ecology and Sociobiology 54 :511-520, 2003
- Seeley T. et Visscher P. Quorum sensing during nest-site selection by honeybee swarms. Behavioral Ecology and Sociobiology 56 : 594-601, 2004.
- Gilley D. et Tarpy D. Three mechanisms of queen elimination in swarming honey bee colonies. Apidologie 36 : 461-471, 2005.
- Kirchner W. Acoustical communication in honeybees. Apidologie 24 : 297-307, 1993.
- Michelsen A. et co-auteurs. The tooting and quacking vibration signals of honeybee queens : a quantitative analysis. Journal of Comparative Physiology A 158 : 605-611, 1986.
- Ramsey M. et co-auteurs. Extensive vibrational characterisation and long-term monitoring of honeybee dorso-ventral abdominal vibration signals. Scientific Reports 8 : 14571, 2018.


